تازه های بیوتکنولوژی
جدیدترین دستاوردهای بیوتکنولوژی، نانوبیوتکنولوژی و بیوانفورماتیکتازه های بیوتکنولوژی
جدیدترین دستاوردهای بیوتکنولوژی، نانوبیوتکنولوژی و بیوانفورماتیکدرباره وبلاگ
 مطالب این وبلاگ جهت معرفی علم بیوتکنولوژی و دستاوردهای جدید آن در زمینه های دارویی، پزشکی، صنعتی و کشاورزی گردآوری شده است و هدف آن اطلاع رسانی به دانشجویان و محققین علاقمند به بیوتکنولوژی و زیست شناسی سلولی مولکولی می باشد. امید است مطالب مورد توجه این عزیزان قرار گرفته و مفید واقع شوند. جهت ارتقای کیفی و علمی وبلاگ ما را از انتقادات و پیشنهادات خود بی بهره نگذارید.
با سپاس فراوان...
آدرس صفحه اینستاگرام: biotech.world@
سیفی
ادامه...
مطالب این وبلاگ جهت معرفی علم بیوتکنولوژی و دستاوردهای جدید آن در زمینه های دارویی، پزشکی، صنعتی و کشاورزی گردآوری شده است و هدف آن اطلاع رسانی به دانشجویان و محققین علاقمند به بیوتکنولوژی و زیست شناسی سلولی مولکولی می باشد. امید است مطالب مورد توجه این عزیزان قرار گرفته و مفید واقع شوند. جهت ارتقای کیفی و علمی وبلاگ ما را از انتقادات و پیشنهادات خود بی بهره نگذارید.
با سپاس فراوان...
آدرس صفحه اینستاگرام: biotech.world@
سیفی
ادامه...
مکانیسم های مولکولی مقاومت به ویروس های گیاهی (مقاله تخصصی)
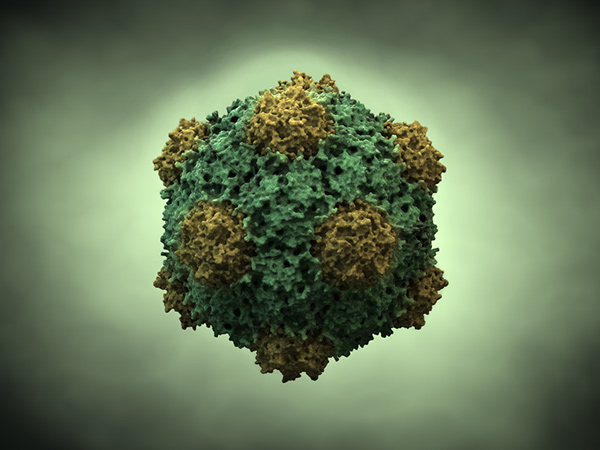
ویروسهای گیاهی ویروسهایی هستند که گیاهان را آلوده میکنند. مشابه تمام ویروسها، ویروسهای گیاهی پارازیتهای اجباری درون سلولی هستند که مکانیسم مولکولی مورد نیاز برای همانند سازی را بدون میزبان ندارند. کشف ویروسهای بیماریزای گیاهی به سال 1898 توسط شخصی به نام مارتینوس بر میگردد. وی با فیلتر کردن برگهایی که علایم موزاییک توتون را نشان میدادند متوجه شد که عوامل دیگری بجز باکتریها که از فیلتر عبور نمیکنند، از فیلتر عبور کرده و میتواند سبب ایجاد بیماری در گیاهان سالم شود. بعد از کشفهای اولیه در مورد مفهوم ویروس و بعد از اختراع میکروسکوپ الکترونی و با ااستفاده از آن تعداد بیشتری از ویروسها شناسایی و در سال 1939 اولین لیست طبقهبندی شده از 129 ویروس گیاهی منتشر شد.
امروزه بیشتر تحقیقات بر روی فهم ژنتیکی و بیولوژی مولکولی ژنوم ویروسهای گیاهی متمرکز شده است. هدف از این تحقیقات فهم چگونگی همانندسازی ویروس، انتقال آنها و آلودگی در گیاهان است. فهم ژنتیک ویروسها و نقش پروتئینهای آنها برای بررسی بنیهی آنها برای استفادهی تجاری در شرکتهای بیوتکنولوژی بوده است. بویژه، توالیهای حاصله از ویروس برای درک بهتر اشکال جدید مقاومت استفاده شده است. همین پیشرفتهای خیره کنندهی سالهای اخیر به محققین این اجازه را داده است تا ویروسهای گیاهی را برای راهکارهای جدید برای تولید پروتئینهای جدید با ارزش به خدمت بگیرد.
مدیریت ویروسهای گیاهی:
راهکارهای متداول برای برای مدیریت بیماریهای ویروسی به طور معمول به صورت زیر تعریف میشود.
الف) مبارزهی شیمیایی که بدلیل هزینهی بالا و اثرات سوء زیست محیطی اگرچه جزو یکی از معمولترین روشهای مبارزه است اما زیاد کارا به نظر نمیرسد.
ب) مدیریت کلاسیک بیماریهای ویروسی با کنترل جمعیت ناقل با استفاده از حشره کشها، استفاده از مواد گیاهی عاری از ویروس، استفاده از کولتیوارهای مقاوم، غربال موتانتها، تلاقیهای برگشتی برای انتقال ژنهای مقاومت از گیاهان مقاوم به حساس، روشهای کشت اختصاصی و ... صورت میگیرد. برای مثال از روش تلاقی برگشتی برای هرمی کردن ژنهای مقاومت به ویروس موزایک سویا حاصل شده است.
ج) تولید ارقام مقاوم که کمهزینهترین و نیز سالمترین روش مبارزه برای ویروسهای گیاهی است. برای مثال در گیاه زینتی گل ساعتی و گیاه زراعی جو انتقال ژنهای مقاومت سبب کاهش و یا حذف علایم ویروس موزایک لوبیا و ویروس ... به ترتیب از روی گیاهان گل ساعتی و جو شده است.
مقاومت مولکولی به ویروسها:
به دلیل اندازه نسبتاً کوچک ژنوم ویروسهای گیاهی توسعه راهکارهای مولکولی برای کنترل بیماریهای ویروسی گیاهی به طور خاص موفق بوده است. راهکارهای متفاوتی برای استفاده از فناوری مولکولی به منظور تلفیق با ایجاد فاکتورهای مقاومتی جدید در سیستمهای ویروسی گیاهان وجود دارد. روال کار به این نحو است که آن دسته از محصولات ژن یا ژنهای ویروسی شناسایی شوند که وقتی در زمان نامناسب یا به مقدار نادرست وجود دارند، با کارکردهای نرمال فرآیند آلودگی دخالت نموده و مانع از پیشرفت بیماری میشوند.
چند نکته قابل تامل در مورد مقاومت:
 گیاه مقاوم گیاهی است که آلودگی را نشان نمیدهد یا آلودگی با نسبت کمتر
یا علایم بیماری با سرعت کمتریی نسبت به گیاهان حساس مشاهده میشود.
گیاه مقاوم گیاهی است که آلودگی را نشان نمیدهد یا آلودگی با نسبت کمتر
یا علایم بیماری با سرعت کمتریی نسبت به گیاهان حساس مشاهده میشود.- آزمایشات مقاومت بر این پایه استوار است که چگونه یک گیاه میتواند پروسهی بیماری را متوقف یا کند سازد.
-
مقاومت به ویروس شامل تداخل در هر مرحلهای از سیکل آلودگی ویروس مثل وارد
شدن به درون میزبان، همانندسازی، حرکت در بین یا در درون سلولها و انتقال
از محل اولیهی آلودگی به بخشهای دیگر گیاه میباشد.
-
اکثر ویروسهای گیاهی ژنوم RNA تک رشتهای مثبت sense دارند اگرچه بعضی از
ویروسهای مهم نیز ژنوم DNA منفی یا مثبت و تک رشتهای یا دو رشتهای
دارند.
جایگاه نشانگرهای مولکولی:
استفاده از نشانگرهای مولکولی در جهت شناسایی ژنهای مقاومت و استفاده از آنها برای انتخاب گیاهان مقاوم در شرایط درون شیشه است. برای شناسایی نشانگرهای پیوسته با ژن مقاومت مراحل زیر به ترتیب انجام میشود. ابتدا باید جمعیت در حال تفرق با زمینهی ژنتیکی یکسان تهیه شود سپس تلقیح عامل بیماریزا به این جمعیت صورت میگیرد. بعد از آن کلیهی افراد جامعه نسبت به مقاومت یا حساسیت به عامل بیماریزا به دو گروه مقاوم و حساس طبقه بندی میشوند. سپس مقایسهی تودهها با والدین حساس و مقاوم صورت گرفته و نشانگرهایی که معرف حساسیت یا مقاومت هستند را میتوان بدست آورد. در انتها تجزیه پیوستگی روی باندها انجام و در صورتی که مکان ژنی نشانگر نزدیک به ژن مقاومت یا حساسیت باشد میتوان از وجود نشانگر به مقاومت پی برد.
نشانگرهای مولکولی ابزاری ارزشمند برای شناسایی تعداد ژنهای مقاومت، مطالعهی رفتار این ژنها و شناسایی محل قرارگیری این ژنهاست. همچنین با استفاده از نشانگرهای مولکولی، نیاز به آزمایش تعداد زیاد بوته در شرایط گلخانه و یا مزرعه تحت شرایط آلودگی کاهش مییابد. امکان شناسایی مقاومت در مراحل اولیه رشد وجود دارد و زمان تولید واریتههای جدید به طور قابل ملاحظهای کاهش مییابد.
مکانیسمهای طبیعی گیاهان برای مقاومت به ویروسها:
مهمترین عامل ساختمانی در ممانعت از ورود ویروسهای گیاهی، ساختار دیوارههای سلولی گیاهی است. دیوارهی سلولی به عنوان ساختار حیاتی محکم به عنوان یک سد طبیعی مانع از ورود اولیه ویروس بدرون گیاه میشود.
مکانیسم دفاع فعال با واکنش فوق حساسیت یکی از مهمترین و موثرترین مکانیزمهای دفاع فیزیولوژیکی است. مرگ سلولهای اطراف آلودگی بر طبق مرگ برنامهریزی شدهی سلول که با ظهور علایم نکروتیک مشخص میشود مانع از انتقال ویروسها به سایر بخشهای گیاه میشود. ایجاد این واکنش بوسیلهی شناسایی اختصاصی ویروس توسط ژنهای غالب مقاومت (R ) به ژنهای ویروسی صورت میگیرد.
مخافظت از آلودگی ویروسی میتواند بوسیلهی ژنهای مقاومت موجود در ژرم پلاسم ها بدست آید. این ژنها میتوانند به صورت غالب R و هم به صورت مغلوب باشند. مثالی از ژنهای R، ژن مقاومت به ویروس Y سیب زمینی، Rx2 و Rx1 برای ویروس X سیب زمینی(PVX)، SW5 برای مقاومت به ویروس پژمردگی لکهای گوجه فرنگی(TSWV) است. برخی از این ژنهای مقاومت R برای ایجاد مقاومت به گیاهان تراریخت منتقل شدهاند. به جز ژنهای R، ژنهای مغلوب طبیعی نیز گزارش شدهاند. مشخص شده است که چندین ژن گیاهی مثل فاکتور شروع یوکاریوتی 4E (eIF4Eiso) برای تکمیل چرخهی آلودگی ویروس در سلولهای گیاهی نیاز به این فاکتور دارند و گیاهان جهش یافتهای که فافد ژن عملکردی eIF4Eiso هستند نسبت به پوتیویروسها مقاوماند.
مقاومت منتج شده از پروتئینهای ساختمانی
مقاومت منتج شده از پروتئین پوششی همسان :
این نوع مقاومت اشاره به مقاومتی دارد که در آن پوشش پروتئینی یک ویروس سبب مقاومت به آلودگی در برابر همان ویروس می شود. این نوع مقاومت برای اولین بار توسط آبل در سال 1986 با بیان ژن پروتئین پوششی ویروس موزاییک تنباکو در گیاه تنباکو نشان داده شد. گیاهان تراریخت حاصله یا علایم آلودگی به ویروس را نشان ندادند یا آنهایی که این علایم را نشان دادند با تأخیر نسبت به نمونههای شاهد علایم آلودگی را ظاهر کردند.
نظریه متفاوتی برای توضیح مکانیسم مقاومت به واسطه پروتئین های پوششی پیشنهاد شده است. مهمترین و مقبول ترین مدل ارائه شده پیشنهاد میکند که گیاهان تراریخته با ژن پروتئین پوششی باعث ممانعت از جزء جزء شدن ویروسهای کامل برای در معرض قرار گرفتن مواد ژنتیکی برای رونویسی می شوند. جزءجزء شدن ویروس یکی از مراحل اولیه آلودگی ویروسی است. که در طی آن انتهای RNA /5 ویروسی در معرض ریبوزوم قرار گرفته و از رویORF آن پروتئین پوششی ترجمه میشود( البته پروتئین پوششی یکی از پروتئینهایی است که ترجمه میشود). عمل ترجمه چند دقیقه بعد از آلودگی سلولهای توتون با ویروس موزائیک توتون (TMV) در پروتوپلاست رخ میدهد. سلولهای آلوده حاوی استریپوزوم، ویرویون های بدون پوشش جزءجزء شده، ریبوزومها و ملکولهای پلی پپتیدی در حال تشکیل هستند. به هر حال در پروتوپلاست سلولهای گیاهی تراریخت با ژن پروتئین پوششی TMV از جزء جزء شدن ویریونها ممانعت به عمل آمده بود. حذف کامل و جزیی پروتئینهای پوششی بوسیله تیمار با آلکالین توانست اثر محافظتی مقاومت بواسطهی پروتیئنهای پوششی را از بین ببرد این نتایج پیشنهاد کرد وقتی ویروس مهاجم انتهای RNA /5 کد کننده زیر واحد پروتئین پوششی خود را آزاد میکند، پروتئینهای پوششی نو ترکیب بلافاصله اجزاء ویروسی را دوباره پوششدار میکنند و بدین طریق از آلودگیشان ممانعت به عمل میآید. از این نتایج بر میآید که CO-translational disassembly در مراحل اولیه آلودگی ویروس مرحلهای کلیدی برای تعیین این مسئله است که آیا ویروس می تواند به مقاومت با واسطه پروتئین پوششی غلبه کند یا خیر؟
جزءجزء شدن ویروس موزائیک تنباکو به واسطه اثرات متقابل واکنشهای تدافعی بین گروه های کربوکسیل - کربوکسیل زیر واحدهای آمینواسیدی مستقر شده در حد فاصل زیر واحدهای پروتئین پوششی استوار است. تغییرات در، ph غلظت یون کلسیم در محیط سلولی می تواند واکنش این گروههای باردار منفی را تسهیل کند و در نتیجه آن ویریونهای بی ثبات شروع به جزءجزء شدن میکنند. به همین خاطر در گیاهان تراریخته با ژن پروتئین پوششی TMV فرض می شود که محیط عمومی متداول، مناسب جهت سرهم کردن ویریونهاست تا جزءجزء کردن آنها .
این توانایی خود سرهم شدن پروتئین پوششی که برای مقاومت بواسطه پروتئین پوششی در TMV ضروری است برای همه ویروسهای گیاهی ضروری نمیباشد. برای مثال بیان تراریخت یک موتانت درTMV که فاقد ویژگی سرهم شدن قطعات ویروسی به صورت خود به خودی بود، هیچ گونه مقاومتی علیهTMV در توتون ایجاد نکرد. شایان ذکر است که در برخی موارد حتی بیان پروتئین پوششی ویروس، آلودگی را افزایش داده است. برای مثال این مسئله در مورد ویروسRYMV در برنج گزارش شده است.
مقاومت به واسطه پروتئین های پوششی ناهمسان :
اندرسون و همکاران در سال 1998 گزارش کردند که تنباکوهای تراریخت که پروتئین پوششی TMV را در سلول های خود جمع می کردند در آلودگی به ویروسهای ناهمسان مثلPVX تاخیر نشان میدهند. این پدیده مقاومت به واسطه پروتئینهای پوششی ناهمسان را نشان میدهد. این نوع مقاومت اغلب سطوح پائینی از مقاومت را نشان میدهد. دیده شده است که این مقاومت به وسیله تداخل کوتاه با گسترش سیستمیک و پیشرفت بیماری هنگامی که مقاومت حاصله از پروتئین پوششی همسان کاهش یافت یا آلودگی سیستمیک نبود، بروز یافت .
سه مدل ممکن برای مقاومت بواسطه پروتئین پوششی ناهمسان پیشنهاد شده است:
الف)پروتئین پوششی نوترکیب اثر متقابل دارد و در دسترس بودن جدایههای RNA های ویروسی ناهمسان برای رپلیکاز محدود میشود. این مدل نمونهای برای پوشش پروتئینی TMV است که می تواند RNA های ویروسی ناهمسان را پوششدار کند.
ب)پروتئین پوششی نوترکیب با یکی یا تعدادی از گیرندههای اصلی مورد نیاز برای آلودگی ویروس و همانندسازی آن تداخل ایجاد میکند. باید توجه داشت که مقاومت به واسطه پروتئین پوششی ناهمسان بخاطر اختصاصیبودن گیرندهها به همان موثری مقاومت به واسطه پروتئین های پوششی همسان نیست.
ج)مقاومت شاید در نتیجه راه اندازی پاسخ های دفاعی درونی میزبان بوسیله پروتئین پوششی نوترکیب باشد.
در زیر نمونه هایی از مقاومتهای ایجاد شده بوسیل هی پروتئنهای پوششی آورده شده است.
مقاومت به واسطه پپتید :
به نظر میرسد کاربرد عمومی مقاومت بواسطه پپتید زیاد گسترده نباشد زیرا نیاز به سطوح بالایی از پروتئینهای نوترکیب می باشد. سطوح بالایی از mRNA نوترکیب لازم است تا بتواند خاموشی ژن بعد از رونویسی راهاندازی شود. به همین خاطر بهبود این راهکار وابسته به بهبود کارایی رونویسی mRNA نوترکیب و ثبات سطح بیان پروتئین است.
اخیرأ رادولف و کالوگوس مقاومت به واسطه پپتید را برای طیف وسیعی از گیاهان در برابر توپسو ویروسها ایجاد کردهاند. غربال آزمایشگاهی بیان پپتید بوسیله سیستم هیبرید دوگانه مخمر آزمایش شد. ابتدا آنها پپتیدهایی را شناسایی کردند که میتوانست با میل ترکیبی به دومین عملکردی نوکلئوپروتئین ویروس TSW متصل شود. همچنین چندین پپتید تشخیص داده شد که میتوانستند به نوکلئوپروتئین ویروسهای نزدیک به خانواده توپسوویروسها متصل شوند. نو ترکیبکردن با این پپتید سبب ایجاد مقاومت گیاهان نوترکیب به TSWV، GRSV و CSNV شد. چون مکانیسم مقاومت به واسطه پپتید وابسته به دخالت غالب منفی نقشهای پروتئینهای ویروسی ضروری است جای تعجب نیست که این فناوری بتواند برای ویروسهای DNA نیز به همین خوبی توسعه یابد. در همین راستا لوپز و اُچا نشان دادند که بیان یک پپتید، رپلیکازTGMV را هدف میگیرد که این مسئله می تواند باعث مقاومت به این جمنی ویروس شود.
مقاومت بواسطه ی پروتئینهای غیر ساختمانی Non-structural protein mediated resistance
ویروسها، پروتئینهای غیر ساختمانی کد میکنند که برای همانندسازی ضروری میباشند. اخیرا تعدادی از این پروتئینهای غیر ساختمانی Replicase کشف شده است که وقتی در گیاهان تراریخت بیان میشوند، درجات بالایی از مقاومت به آلودگی ویروسی را ایجاد مینمایند. گلمبوسکی و همکاران در سال 1990 برای اولین بار این پدیده را با بیان چهارچوب قرائت باز 54 کیلو دالتونی ویروس TMV در توتون تراریخت نشان دادند. توتون تراریخت مقاوم به قهوهای شدن زودرس PEBV و سیب زمینی مقاوم به ویروس X (PVX) تولید شد.
مقاومت به واسطه RNA
خاموشی ژن بعد از رونویسی PTGS
خاموشی ژن بعد از رو نویسی یک مکانیسم محافظت شده برای تنظیم mRNA در گیاهان، حیوانات و قارچهاست. این مکانیسم به وسیله تجزیه ذاتی یا تراریخت mRNAها در سیتوپلاسم مشخص میشود که در نتیجه آن بیان ژن کاهش مییابد. در گیاهان این نوع خاموشی ژن فرآیندهای نموی متعددی را کنترل میکند و تجمع آن تنظیم کنندهی ایمنی ذاتی به ویروسهاست. دو نشانه برای خاموشی ژن بعد از رونویسی وجود دارد . اول اینکه خاموشی mRNAهای هدف در سیتوپلام رخ میدهد و دوم مولکولهای کوچک RNA (21-25 نوکلئوتیدی) از mRNAهای هدف خاموش شده به وجود میآید. دو نوع RNA کوچک با توجه به تفاوتهایشان در بیوژنز به نام siRNAو miRNA تشخیص داده شدهاند. اینکه چگونه مسیرهای siRNA و miRNA مینوانند در ایجاد مقاومت استفاده شوند در زیر بحث می شود.
الف) مسیرsiRNA :
در مسیرsiRNA ، خاموشی ژن بعد از رونویسی بوسیله RNA های ویروسی تک رشتهای که بسیار ساخته شدهاند و یاRNA های دو رشتهای که توسط میزبان یا RdRp(RNA پلی مرازهای وابسته به RNA) تولید شده آغاز میشود. در گیاهانRNA پلی مراز I وابسته به RNA و RDR6 برای خاموشی ویروسها و تراریختها لازم است. اینRDR پلی مرازها ازRNA های تک رشتهای به عنوان الگو برای تولیدRNA های دو رشتهای استفاده میکنند. اینRNA های دو رشتهای به عنوان سوبسترا برای دایسر (یک نوع III Rnase) بکار می رود. دایسر میتواند RNA دو رشتهای را بهsiRNA های با طول 21-25 نوکلئوتید برش دهد. siRNAهای تولید شده با مجموعههای خاموشی القاء شونده باRNA (RISC) ادغام میشوند که در نهایت برشRNA های هدف صورت میگیرد. به صورت متناوب این مجموعهها میتوانند به درون مجموعههای خاموشکننده رونویسی که باRNA القا میشوند وارد شده تا کروماتین را بطور مستقیم از طریق متیلا سیون هیستون وDNA خاموش کنند. در مجموعه RISC , siRNA در میان اتصال شونده خاص توالی محل برشRNA های هدف واقع میشوند. قطعات برش یافته در سیتوپلاسم توسط اگزونوکلئازها به طور کامل تجزیه می شوند. به طور متناوب siRNAها به عنوان پرایمر برایRDR پلی مرازها برای رو نویسیRNAهای هدف مولد RNAهای دو رشتهای و siRNAهای بیشتر استفاده میشود. این RDR پلی مرازها فعالیت خزانه siRNAها را افزایش و در نتیجه آن خاموشی افزایش مییابد.
آرابیدوپسیس حاوی چهار پروتئین دایسر (DSL) است. سه تا از آنها در مسیر siRNA هنگامی که DCL1 برای مسیرmiRNA لازم است، شرکت دارد. 2 DCL رونوشت آنتی سنس طبیعی مرتبط با تنش را سنتنز میکند، DCL3 ، siRNA بیست و چهار نوکلئوتیدی درگیر در متیلاسیون و شکلگیری هتروکروماتین را تولید میکند و 4DSL ، siRNAفعال ترانس 21 نوکلئوتیدی، که حد فاصل خاموشی ژن بعد از رونویسی برخی از ژنهای درونزاد یا ترانسژن است را تولید می کند .
2 DCLو4DSL در تولید siRNA از برخی ویروسها مثل TCV، CALCUV ، CMV،TRV درگیرند. بنابر مدارک اخیر پیشنهاد میکند که پروتئینهای DCL نقش همپوشان و قابل تعویض در هر دو مسیر siRNA و miRNA دارند .
مقاومت با واسطه RNA در ویروس هایRNA و DNA:
مقاومت با واسطه RNA میتواند مقاومت علیه گسترده وسیعی از ویروسهایRNA و DNAرا ایجاد کند. در مقایسه با مقاومت بواسطه پروتئین پوششی، مقاومت بواسطهRNA سطح بالاتری از مقاومت به ویروس را ایجاد می کند. به هر حال این نوع از مقاومت تنها علیه ویروسهایی که توالی آنها رابطه نزدیکی با هم دارند ظاهر میشود. و این مسئله به عنوان یک نقص برای مقاومت به ویروس در محصولات زراعی مطرح است. آلودگی توسط چند ویروس به طور معمول در شرایط مزرعهای رخ میدهد. آلودگی با گونههای ویروسی با توالی غیر مرتبط ممکن است بر مقاومت به واسطه RNA غلبه کند چون خاموشی ژن پس از رونویسی بوسیلهی بازدارندههای خاموشی از ویروسهای غیر مرتبط کاهش یافته یا به کلی متوقف میشود. بنا بر این کار برد عملی این مقاومت نیاز به توسعه راهکار هایی دارد که مقاومت ویروسی به طیف وسیعی از ویروسها در مزرعه را ایجاد کنند.
جان و همکاران در سال 2000 سازهای ژنتیکی ساده با توالیهای بدست آمده از دو ویروس مشخص (TGMV، TSWV) ساختند. بیان تراریخت این ژنها توانست مقاومت بواسطه RNA علیه چندین ویروس ایجاد کند. به طور مشابه بوچر و همکاران 2006 نشان دادند که گیاهان گوجه فرنگی بیان کننده یک RNA سنجاقسری شمیریک ساده و کوچک که شامل چهار قطعه ژنی توپسوویروس بود (TSWV، GRS ،TCSV ،WSMOV) مقاومت علیه طیف وسیعی از توپسوویروسها را نشان داد.
مقاومت بواسطه RNA به عنوان یک راهکار موثر علیه ویروسهای DNA جمنی ویروسها مطرح شده است. ایبر (2007) دریافت که بیان ترانسژن siRNA جمنیویروس قبل از آلودگی ویروس حتما باعث ایجاد مقاومت به ویروس نمی شود. این مشاهدات وجود یک آستانه بیان siRNA قبل از اینکه ویروس از پایش و تولید حجم بحرانی ویروسها ( که در سراسر گیاه پخش خواهند شد ) فرار کند را پیشنهاد کرد.
از طرف دیگر بیان (2006) نشان داد که جمنی ویروسها میتوانند از مقاومت به واسطه RNA به خاطر همانندسازی اپی زومی فرار کنند. این دانشمند سطوح بالای متیلاسیون در DNA تراریخت و DNA ویروسی را در گیاهان آلوده با جمنی ویروسها مشاهده کرد.
به هر حال اثر خاموشی نمی تواند همانندسازی جمنی ویروس ممانعت کند به همین خاطر این رویکرد مقاومت به واسطه RNA نمی تواند بر روی ویروسهای DNA دارکارآ باشد.
مسیر میکرو RNA (miRNA) :
miRNA ها RNA های تک رشتهای با 21 نوکلئوتید طول هستند که از پیرایش پیش ساز miRNA بوسیله دایسر بوجود میآید. تجزیه تحلیل پیش سازهای miRNA مشخص کرده است که برای پیرایش صحیح توسط دایسر نیاز به یک ساختار صحیح ثانویه است . و به طور نمونه، پیش سازهای miRNA بواسطه جفت نشدن کامل (mismatching) توالی دارای ساختارهای شبهه سنجاق سری با حلقه در ساقه میشود. مفهوم این جفت نشدن هنوز مشخص نیست اگر چه این جفت شدن ممکن است برای بیوژتر miRNA صحیح هنگامی که آنها میخواهند در راهنمایی دایسر به پیرایش صحیح miRNA 21 نوکلئوتیدی درگیر باشند، لازم باشد.
در پرتروتوزوآها، رونوشت اولیه miRNA (miRNA pri-) چندین کیلو باز طول دارد و حاوی 5-cap و دنباله پلی آدنین است . این رونوشت طویل بوسیله درشا (drosha) که یک ریبو نوکلئاز نوع III است پردازش میشود. این پردازش با کوفاکتور ویژه آن DGCR8 حاوی دومین متصل شونده به RNA دو رشتهای است که میتواند محل اتصال RNA دو رشته ای و تک رشته ای را در رو نوشت اولیه miRNA (pri-miRNA) شناسایی کند و بنابراین drosha را برای پردازش دقیق رو نوشت اولیه و آزاد سازی یک قطعه 65 نوکلئوتیدی شبه سنجاق سر، پیش ساز miRNA (miRNA pri-) هدایت کند. بعد از این که premiRNA یا پیشساز miRNA بوسیله مجموعه 5-ran-GPT اکسپورتین به سیتوپلاسم منتقل شد بوسیله دایسر به miRNA بالغ تبدیل می شود.
سیر تکاملی miRNA در آرابیدوپسیس به طور کاملأ محافظت شده نیست زیرا ژن های کد کننده همو لوگ drosha و DGCR8 بوسیله یک همولوگ دایسر به نام پروتئین هسته ایDCL1 مهیا میشود. DCL1 قادر است که پیش ساز miRNA را به miRNA بالغ پردازش کند. miRNA بالغ در سیتوسول به مجموعه RISC اضافه می شود. تاکنون 184 ژن miRNA در آرابیدوپسیس تالیانا تشخیص داده شده و چندین ژن از آنها تنظیم فعالیتهای نموی مهم گیاه را نشان دادهاند.
مقاومت به واسطه miRNA های مصنوعی (amiRNA) :
گزارشات نشان داده اند که تغییر چندین نوکلئوتیدی از توالی miRNA بیست و یک نوکلئوتیدی اثری بر روی سیر تکاملی و بلوغ آن ندارد. این یافتهها امکان طراحی دوباره توالی miRNA برای رو نوشتهای خاص هدف که به طور ذاتی تحت کنترل miRNA نیستند را بوجود آورد. نیو وهمکاران در سال 2006 کاربرد بیوتکنولوژیایی، تکنولوژی amiRNA را در تحقیق درباره مقاومت ویروسهای گیاهی نشان داند. پیش سازpri-miR159a برای تولید دو amiRNA که مکمل توانایی های ژنوم های دو ویروس گیاهی بودند، استفاده شد. (TUMV , TYMV) وقتی این دو ژن در سطح مناسبی بیان شدند مقاومت اختصاصی به هر دو ویروس بسته به بیان amiRNA همجنس مشاهده شد.
مقاومت به چند ویروس و غلبه بر اثر ات محیطی amiRNA :
در مزرعه آلودگی با چندین ویروس می تواند کارآیی ضد ویروسی را در محصولات زراعی تراریخت به خطراندازد. مثلا متوقف کنندههای ویروسی بیان شده از گونه های ویروسی دیگر می توانند مقاومت به واسطه amiRNA یا siRNA را ازبین ببرند. بیان همزمان دو پیش ساز amiRNA در یک رو نوشت برای تولید پیش ساز دوگانه amiRNA میتواند برای تولید دو amiRNA و ایجاد مقاومت به دو ویروس مختلف استفاده شود. بنابراین راهکار amiRNA چندگانه می تواند به طور وسیع برای بهبود آلودگی های چند گانه در مزرعه به کار رود.
شرایط محیطی مثل رطوبت کم می تواند بر مقاومت بواسطه siRNA تاثیر گذارد. همچنین گزارش شده است که ویروس و خاموشی RNA بواسطه انتقال ژن در رطوبت پایین کاهش داشته است و علت آن ممانعت از تجمع siRNA در سلول های گیاه، حشره، پستانداران در این شرایط بوده است. این حساسیت به رطوبت شکستن مقاومت به ویروس به واسطه siRNA درC 15ْ را توضیح می دهد. از طرف دیگر تجمع amiRNA به سختی در رطوبت پایین تحت تاثیر قرار میگیرد و لاین نو ترکیب بیان کننده amiRNA ها مقاومت ویروسی اختصاصیشان را حتی در رطوبت پایین حفظ کردند.
محافظت متقاطع :
علاوه بر مقاومت بواسطه پروتئین وRNA ، محافظت متقاطع برای کنترل بیماریهای ویروسی استفاده شده است. این مقاومت در یک گیاه میزبان بوسیله آلودگی با یک نژاد ضعیف (ویروس های محافظتی ) یک ویروس القاء میشود. در نتیجهی این آلودگی، گیاهان آلوده در آلودگیهای بعدی به نژاد های مرتبط نزدیک با ویروس مقاومت میکنند. ویروسهای محافظتی نقطه کلیدی برای محافظت متقاطع ویروسهای بیماریزا هستند. ویروسهای محافظتی، ویروسهای ضعیف شدهای هستند که سبب ایجاد علایم خفیف یا بدون ایجاد علایم بیماری در گیاه هستند. یک بار آلودگی با نژاد ضعیف ویروس می تواند سبب ایجاد مقاومت در برابر ویروسهای بیماریزا را به همراه داشته باشد. برخی از دانشمندان یافتهاند که جهش در ممانعت کنندههای خاموشی و ویرسی می تواند علایم جدی را کاهش دهد. مثال آن جهش در ممانعت کنندههای خاموشی ویروسی HC-PRO از پتی ویروس هاست. همچنین موتاسیون در قطعه 126 دالتونی رپلیکاز، دیگر ممانعت کننده خاموشی از ویروس PMMOV منجر به تقلیل علایم و ایجاد مقاومت متقاطع در گیاهان فلفل شد.
یک ویروس رشته منفی مهم، ویروس پژمردگی لکهای گوجه فرنگی (TSWV) است. در این ویروس، RNA ژنومی شدیداً با پروتئین نوکلئو کپسید پیوند خورده است. این پروتئین در بسته بندی RNA ویروسی و همچنین در بسته بندی RNA ویروسی و همچنین در تنظیم مراحل رونویسی تا همانند سازی در طی سیکل آلودگی، فعالیت مینماید. با استفاده از این روش گیاهان تراریخت در توتون و گوجه فرنگی تولید شدهاند.
پدیده محافظت متقاطع بسیار شبیه به پاسخهای ایمنی در جانوران است. پاسخی که بوسیله واکسن ویروسی ضعف شده القاء میشود. به هر حال گیاهان دارای چنین سیستم مشابهی نیستند و مکانیسم حفاظت متقاطع روشن نشده است. این مقاومت ممکن است بوسیله پروتئین،RNA و یا ترکیبی از هر دو مکانیسم باشد.
محافظت متقاطع بوسیله پروتئین :
نقش پروتئینهای پوششی در پاسخهای محافظت متقاطع در آزمایشات متفاوتی تجزیه و تحلیل شده اند.
1)گیاه N.sylvesteris با ویروس موزاییک تنباکو به طور سیستمیک آلوده شد و نواحی سبز تیره و روشن در برگها رشد یافت. (نواحی موزاییکی در پس زمینه سبز برگ ) نواحی موزاییکی سبز روشن حاوی مقادیر بالاتری از TMV علیه مایهکوبی با یک نژاد بیماریزای TMV که نکروزه کننده برگ بود مقاومت نشان میداد در حالیکه منطقه سبز تیره حاوی مقادیر کمتری از ویروس TMV بود ونسبت به آن نژاد نکروزه کننده TMV مستعدتر بود. این نتایج نشان داد که حضور نژاد محافظتی برای مقاومت ضروری است. از طرف دیگر این مقاومت می تواند بوسیله آلودگی باRNA ویروسی بدون پوشش از بین برود.
2)نژاد بدون پروتئین پوششی TMV به نام DT-1G هیچ گونه محافظتی علیه نژاد نکروز کننده TMV ایجاد نکرد.
3)مشابه آنچه در مقاومت مرتبط با پروتئین پوششی دیده شد، موتانتهای متفاوت در پروتئین پوششی با افزایش یا کاهش اجتماع پروتئین پوششی ( وقتی وکتور ویروسی PVX به طور سیستمیک بیان شد ) نشان دادند که جهش یافته پروتئین پوششی با قابلیت اجتماع بالا، حفاظت متقاطع مناسبی را علیه آلودگی با TMV بوجود میآورد. بنابراین حفاظت متقاطع TMV و مقاومت مرتبط با پروتئین پوششی میتواند دلیلی بر توانایی پروتئین پوششی در ممانعت از بدون پوشش شدن ویروسهای مشکلزا باشد.
4)دیتریچ و همکاران (2003) توزیع پتی ویروسها را که با فلور سنتهای متفاوتی نشاندار کرده و آنها را در آلودگیهای مخلوط شدهای به گیاهان N. sylvestris انکوبه کرده بودند، بررسی کردند. گیاهان آلوده شده بوسیله هر کدام از مخلوطهای PVX/ PVYS علایم مشترکی با تعداد زیادی از گیاهان آلوده شده مضاعف که هر دو نوع فلورسنت را نشان میدادند، نشان داده شد. بر عکس در مورد نژادهای PVYS که به صورت متفاوت و نشانمند شده بودند توزیع فلورسنت سبز و قرمز به صورت همبارز در سلول های آلوده تفرق نشان داد. این نتایج نشان داد که در آلودگیهای مختلط بوسیله ویروسهای مشابه اولین ویروس از آلودگیهای بعدی ویروس هم نژاد ممانعت میکند در حالی که این موارد برای ویروسهای غیر مشابه صادق نبود. از این رو مکانیسم مقاومت بواسطه پروتئین می تواند برای محافظت متقاطع علیه ویروسهای مشابه استفاده شود. به طور جالب TMV بدون پوشش شده هنوز سبب محافظت متقاطع میشود. که این مسئله میرساند که ممکن است مکانیسمهای جایگزین دیگری که نیازمند محافظت بر پایه RNA هستند وجود داشته باشد.
محافظت متقاطع بواسطهRNA :
برخی از ویروس های گیاهی مثلCAMV ,TBRV ,TRV به عنوان ویروسهای القاء کننده ترمیمی طبقهبندی میشوند. این ویروسها منجر به ایجاد علایمی بر روی برگهای آلوده می شوند اما علایم در برگهای بالایی با گذشت زمان ترمیم می شوند. این ویروسهای تحریک کننده ترمیمی، خاموشی ژن با القاء ویروسی (VIGS ) را هدف قرار داده و محافظت متقاطع علیه نژادهای ویروسی با ارتباط نزدیک یا با توالی مرتبط در آلودگیهای بعدی را به گیاه میبخشد. برای مثالTRV-GFP خاموشی ژن با القاء ویروسی را تحریک کرده و محافظت متقاطع علیه PVX-GFP ایجاد میکنند. و همچنینTBRV میتواند خاموشی ژن با القاء ویروسی را تحریک کرده و محافظت متقاطع علیه PVX-W22 که حامل قطعهای از ژنوم TBRVاست، بوجود آورد. این یافتهها از این نظریه حمایت میکند که مکانیسم با واسطه RNA رادر محافظت موثر میداند.
به طور جالبی، نژاد ملایم ویروس ZYM به نام GAC که دو جهش در موتیف محافظت شده HC-PRO دارد علایم فتوتیپ ترمیمی را در کدو القاء میکند در حالی که نژادWTZYM سبب بروز علایم شدید بیماری میشود. گیاهان آلوده شده با WT و موتانت GAC هر دو علایم را نشان دادند و تیتر ویروس در 5dpi افزایش یافت. در هر حال بین 5-10 dpiبرگهای سیستمیک گیاهان آلوده شده با GACعلایم بیماری را از دست دادند و تیتر GACکاهش یافت در حالی که تیتر نژاد WT در 5dpi به سرعت افزایش یافت و سپس به یک سطح ثابت رسید این مسئله نشان داد که کارایی محافظت متقاطع با روز های بعد از آلودگی ویروس محافظتی و با تیتر GAC مرتبط است بویژه اگر گیاهانی که با GAC آلوده شده بودند با یک نژاد بیماریزای شدید بعد از 5dpi آلوده میشدند، مقاومت کاملی بدست میآمد، گیاهان آلوده با GAC مقاومتی ناقص یا علایم شیمیریک نشان می دادند.
احتمال دارد که GAC خاموشی ژن با القاء ویروسی را برای ایجاد مقاومت علیه نژاد بیماریزای شدید (WT) هدف قرار داده و جهش یافته HC-Pro نتواند به طور کارآیی از خاموشی ژن با القاء ویروسی ممانعت کند که در نهایت منجر به کاهش تیتر GAC بعد از 5dpi می شود. به طور تعجب آوری، جهش یافته HC-Proدر GAC به طور کامل توانایی خود مبنی بر ممانعت از خاموشی ژن را از دست نداده بود اما وقتی GAC در گیاهان نوترکیب تزریق شد ممانعت از خاموشی ژن با القاء ویروسی ادامه یافت. بنابراین پاسخهای متعددی میتواند در محافظت متقاطع درگیر باشد. که نمیتوانند به سادگی و با یک مکانیسم ساده توضیح داده شوند.
مکانیسم های چند گانه محافظت متقاطع :
مقدار کمی از نژاد های ویروسی ضعیف در محافظت متقاطع گیاهان آلوده شده با GAC حاصل میشود. این RNAهای ویروسی ممکن است هدف قرار گرفته و به طور پیوسته خاموشی ژن با القاء ویروسی را حفظ کرده و مقاومت علیه آلودگی نژاد بیماریزای جدی را ایجاد میکنند. علاوه براین پروتئین پوششی نژاد ضعیف در سلولهای گیاهی وجود دارد و بنابراین ممکن است بدون پوشش شدن نژاد ویروسی ویژگی بیماریزایی شدید را متوقف کنند. ممانعت کنندههای ویروسی جهش یافته مثل HC-Pro ممکن است با چرخه زندگی نژاد ویروسی با ویژگی بیماریزایی شدید بوسیله اثر منفی غالب تداخل ایجاد کند. در این مورد محافظت ناقص، توضیحات تیتر نژاد های ضعیف و قوی اثر دفع متقابل را نشان می دهد. این دفع متقابل پیشنهاد می کند که دو ویروس در آلودگی هم زمان گیاه با هم رقابت میکنند به هر حال این امکان وجود دارد که نژاد ضعیف پاسخ ایمنی اولیه گیاه را با مکانیسمهای ناشناخته دیگری را در گیاه راه اندازی کند. به طور خلاصه محافظت متقاطع بوسیله مجموعهای از مکانیسمهای دفاعی متنوع صورت می گیرد. این مکانیسمها شامل مقاومت بواسطه RNA و پروتئین و همچنین شامل پاسخهای ایمنی گیاه و رقابت ویروس - ویروس میباشد.
حفاظت با RNA ماهوارهای Satellite RNA protection :
RNA های ماهوارهای، گروهی از RNAهای تک رشتهای کوچک( تقریباً 300 نوکلئوتید) هستند که برای همانند سازی و بسته بندی ویریونی به منظور ایجاد آلودگی در جای دیگر به یک ویروس کمکی (Helper virus) وابسته میباشند. بنابر این RNA ماهوارهای برای تکثیر و انتقال به ویروس وابسته است، اگر چه وابسته به ژنوم ویروس نیست. این گونه RNA ماهوارهای با چندین ویروس دیگر مرتبط هستند. تعدادی از RNAی ماهوارهای تکثیر و علایم ویروس کمکی خود را تعدیل مینمایند. بسته به RNAی ماهوارهای مرتبط، طیف تغییر در ایجاد علایم از نکروز شدید تا کم اثر شدن شدید آن میباشد. بنابر این RNAهای ماهوارهای که علایم را تغییر میدهند میتوانند به طور بالقوه برای کاهش شدت بیماری ویروس کمکی مورد استفاده قرار گیرند. بدین جهت کاربرد آن در گیاهان تراریخت برای ایجاد مقاومت در گیاهان زراعی از جایگاه مهمی برخوردار گردیده است. تاین و همکاران در سال 1991 نشان دادند که تلقیح عمدی یک نژاد ویروس موزاییک خیار CMV حاوی یک RNAی ماهوارهای تقلیل دهندهی علایم، گیاهان توتون، فلفل، گوجه فرنگی و خیار را به طور موفقیتآمیزی در برابر یک نژاد بیماریزای CMV محافظت نمود و میزان خسارت را کاهش داد.
تاین و گوسوی در سال 1991 گزارش کردند که 121 گیاه گوجه فرنگی تراریخت بیان کنندهی یک RNA ماهوارهای تقلیل دهندهی علایم CMV، در نقایسه با گیاهانی که یک نژاد قوی CMV به آنخا تزریق شد، 50% عملکرد بیشتری تولید کردند. این راهکار مربوط به آن دسته از سیستمهای ویروسی است که دارای RNAی ماهوارهای تقلیل دهنده هستند. کیم و همکاران فلفلهای قرمزی تولید کردند که RNA ماهوارهای CMV را بیان کردند. کاهش علایم ویروسی در نتاج این گیاهان، پس از تلقیح با نژادهای CMV-Y یا CMV-Korea تایید گردید.
مقاومت بواسطهی سنس (sense) و آنتی سنس (Antisense):
استراتژی دیگر الگو گرفته از پاتوژن که برای کنترل ویروسهای گیاهی مورد بررسی قرار گرفته است، بیان ترانسژن آنتی سنس و جدیداً قطعات سنس RNAهای ویروسی میباشد. اساس این راهکار، اتصال RNA ویروسی با توالیهای RNA مکملی است که توسط گیاه بیان میشود. جفت شدن نامناسب RNA-RNA، از در دسترس بودن RNA ویروسی برای همانند سازی و بیان ژن، ممانعت مینماید. بنابر این ساختارهای آنتیسنس و سنس میتوانند برای بلوکه کردن مراحل اولیه مهم که در ایجاد آلودگی مهم هستند، مورد استفاده قرار گیرند. حفاظت آنتی سنس، در توتون که RNA مکمل پروتئین پوششی ویروس را بیان میکند نشان داده است.
مقاومت به واسطهی پروتئینهای حرکتی Movement protein
پروتئینهای حرکتی برای جا به جایی سلول به سلول ویرویهای گیاهی ضروری هستند. این پروتئینهای حرکتی نقش دریچهی پلاسمودسماتا را تغییر و بدین جهت اجازه انتقال اجزا ویروس یا مشتقات نوکلئوپروتئین به سلولهی مجاور را میدهند.
اساس مقاومت حاصل از پروتئینهای حرکتی بدینگونه است که ار ژنهای جهش یافتهی پروتئین حرکتی برای تولید پروتئین حرکتی با عملکرد نادرست استفاده می-شود در نتیجه این پروتئین ناقص باعث مداخله در فرآیند انتقال و در نهایت باعث تضعیف با تاخیر در آلودگی ویروس میشود. در خقیقت مقاومت حاصله بر اساس رقابت میان دو پروتئین حرکتی ویروس بیماریزا و نوترکیب برای اتصال به جایگاه پلاسمودسماتا میباشد. در گیاهان تنباکو چنین مقاومتی با استفاده از بیان نوترکیب پروتئین حرکتی ناقص TMV در برابر TMV و ویروس موزاییک brome حاصل شد.
منبع:
http://agriculturalbiotechnology.persianblog.ir
سلام .یه سری منابع در مورد انتقال ژن به وسیله ویروس بهم معرفی کنید.