تازه های بیوتکنولوژی
جدیدترین دستاوردهای بیوتکنولوژی، نانوبیوتکنولوژی و بیوانفورماتیکتازه های بیوتکنولوژی
جدیدترین دستاوردهای بیوتکنولوژی، نانوبیوتکنولوژی و بیوانفورماتیکدرباره وبلاگ
 مطالب این وبلاگ جهت معرفی علم بیوتکنولوژی و دستاوردهای جدید آن در زمینه های دارویی، پزشکی، صنعتی و کشاورزی گردآوری شده است و هدف آن اطلاع رسانی به دانشجویان و محققین علاقمند به بیوتکنولوژی و زیست شناسی سلولی مولکولی می باشد. امید است مطالب مورد توجه این عزیزان قرار گرفته و مفید واقع شوند. جهت ارتقای کیفی و علمی وبلاگ ما را از انتقادات و پیشنهادات خود بی بهره نگذارید.
با سپاس فراوان...
آدرس صفحه اینستاگرام: biotech.world@
سیفی
ادامه...
مطالب این وبلاگ جهت معرفی علم بیوتکنولوژی و دستاوردهای جدید آن در زمینه های دارویی، پزشکی، صنعتی و کشاورزی گردآوری شده است و هدف آن اطلاع رسانی به دانشجویان و محققین علاقمند به بیوتکنولوژی و زیست شناسی سلولی مولکولی می باشد. امید است مطالب مورد توجه این عزیزان قرار گرفته و مفید واقع شوند. جهت ارتقای کیفی و علمی وبلاگ ما را از انتقادات و پیشنهادات خود بی بهره نگذارید.
با سپاس فراوان...
آدرس صفحه اینستاگرام: biotech.world@
سیفی
ادامه...
اندونوکلئاز ها
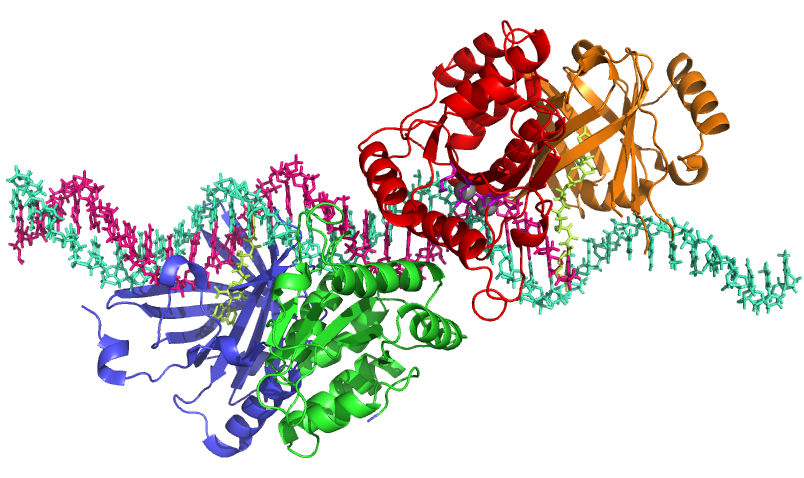
آنزیمهای اندوکلئاز به دو دسته اختصاصی و غیر اختصاصی تقسیم میشوند. دسته اول توالی ویژهای را شناسایی کرده و برش میدهند که اصطلاحا اندونوکلئازهای محدودکننده نامیده میشوند. این نوع آنزیمها از اهمیت ویژهای در کلونینگ DNA بر خوردار میباشند. دسته دوم از این آنزیمها به طور تصادفی اسیدهای نوکلئیک را برش میدهند. در زیر به معرفی تعدادی از آنزیمهای اندونوکلئاز غیر اختصاصی پرداخته میشود.
DNaseI
این آنزیم فقط قطعات دو رشتهای DNA را برش میدهند. از آن در حذف DNA در هنگام خالص سازی RNA، فوت پرینتینگ، شناسایی نواحی فعال کروماتین از نظر نسخه برداری استفاده میشود.
نوکلئاز S۱
این آنزیم از آسپرژیلوس اریزا بدست میآید و در آنالیز هیبریدهای RNA/DNA برای شناسایی اینترون ها، ایجاد مولکولهای دو رشتهای DNA با انتهای صاف و باز کردن ساختارهای سنجاق سر مانند که در هنگام سنتر cDNA ایجاد میشوند بکار میرود. در برخی موارد از آنزیم نوکلئازمونگ بین بجای نوکلئاز S۱ برای این عمل استفاده میشود. آنزیم فوق از نوکلئاز S۱ اختصاصی تر عمل مینماید.
نیکاز
این دسته از آنزیمها فقط یکی از زشتهها را بر روی مولکولهای دو رشتهای DNA برش میدهند. مانند آنزیمهای HincII و gpA. آنزیمgpA بهوسیله فاژ فیX۱۷۴ تولید میشود و در نشاندار کردن DNA به کار برده میشود.
نوکلئاز P۱
این آنزیم از باکتری پنسیلیوم کریزوژنوم بدست میآید و کلیه انواع اسیدهای نوکلئیک اعم از تک رشتهای و دو رشتهای را برش میدهد. از این آنزیم در آزمایشهای مختلف از جمله الکتروفورز با تغییر حرکت (EMSA) کاربرد دارد.
RNaseA
این آنزیم از پانکراس گاو بدست میآید و دارای فعالیت اندونوکلئازی بر روی مولکولهای تک رشتهای و دو رشتهای RNA است. برای حذف RNA در هنگام تلخیص DNA و همچنین شناسایی جهشهای نقطهای در مولکول mRNA استفاده میشود.
RNaseH
این آنزیم رشتههای RNAای که بصورت هیبرید با DNA (RNA/DNA) میباشند را تجزیه میکند. کاربرد آن در تولید cDNA، شناسایی هیبریدهای RNA/DNA میباشد.
RNase ONE
این آنزیم فقط مولکولهای RNA تک رشتهای را برش میدهد و در تعیین توالی RNA، آزمون محافظت در برابر ریبوکلئاز، شناسایی نواحی غیر مکمل در هیبریدهای DNA/RNA کاربرد دارد.
اندونوکلئازهای محدودکننده
اندونوکلئازهای محدودکننده( Restriction Enzyme)، آنزیمهایی هستند که به طور اختصاصی، توالی خاصی را شناسایی کرده و آن را برش میدهند. در صورتیکه توالیهایی مزبور توسط عواملی مثل متیلهشدن تغییر یافته باشند مورد شناسائی آنزیم قرار نمیگیرند. آنزیمهای محدودکننده بر حسب نوع فعالیت و توالی مورد شناسائی به سه دسته تقسیم میشوند. در زیر به معرفی برخی از آنها میپردازیم.
آنزیمهای محدودکننده نوع اول (I)
این دسته از آنزیمها علاوه بر خاصیت برش دهندگی، دارای خاصیت متیلهکنندگی نیز هستند. از جمله این آنزیمها میتوان به آنزیمهای EcoK و EcoB که به ترتیب جایگاههای AACNNNNNNGTGC و TGANNNNNNNNTGCT را شناسایی میکنند. اگر یکی از دو رشته DNA در جایگاه مربوطه متیله باشد این آنزیم رشته مقابل را نیز متیله میکند اما اگر هیچ یک از دو رشته متیله نباشد این آنزیم DNA را در جایگاهی غیر اختصاصی که بیش از ۱۰۰۰ نوکلئوتید از جایگاه شناسایی فاصله دارد برش میدهد. این آنزیم ها به کوفاکتور ATP وS-Adnosyl Methionin احتیاج دارند و ATP را هیدرولیز میکنند.
آنزیمهای محدودکننده نوع دوم (II)
این دسته از آنزیمهای محدودکننده از متداولترین نوع آنزیمهای محدودکننده میباشند و کاربرد وسیعی در کلونینگ ژنها، برش DNA به قطعات کوچکتر، نقشهبرداری فیزیکی ژنوم، ایجاد مولکولهای DNA نوترکیب دارند. این آنزیمها توالیهای ویژهای را روی دو رشته DNA شناسایی کرده و آنرا برش میدهند. جایگاه شناسایی این آنزیمها به صورت واروخوانه (پالیندروم) میباشند. در حالت واروخوانه دو رشته DNA مکمل و معکوس یکدیگر میباشند و دارای نقطه تقارن هستند. در این نوع مولکول DNA از دو طرف َ۳ به َ۵ و َ۵ به َ۳ یکسان خوانده میشود. به طوریکه هر رشته میتواند بر روی خود تا بخورد و ساختار سنجاق سر ایجاد کند. جایگاه شناسایی این دسته از آنزیمها بین ۴ تا ۶ نوکلئوتید و گاهی تا ۸ نوکلئوتید میباشد.
برخی آنزیمهای محدودکننده بدست آمده از منابع متفاوت جایگاههای یکسانی را شناسایی میکنند و آن را به طور یکسان برش میدهند. به چنین آنزیمهائی اصطلاحاً ایزوشیزومر گفته میشود. مثلاً جایگاه شناسائی و محل برش آنزیمهای محدودکننده HpaII و MspI بصورت C/CGG است.
برخی از آنزیمهای محدودکننده جایگاه مشابهی را شناسایی میکنند ولی آن را به دو صورت مختلف برش میدهند، این آنزیمها نئوشیزومر نامیده میشوند. آنزیمهای محدودکننده SmaI و XmaI نئوشیزومر اند و جایگاه و محل برش آنها به ترتیب بصورت CCC/GGG و C/CCGGG میباشد.
برخی اندوکلئازهای محدودکننده که توالی هدف متفاوتی دارند، ممکن است انتهای چسبنده یکسانی ایجاد نمایند. برای مثال، آنزیمهای BamHI با جایگاه شناسائی G/GATCC، BglII با جایگاه شناسائی A/GATCT و Sau۳A با جایگاه شناسائی /GATC، یک نوع انتهای چسبنده ایجاد میکنند. از اینرو انتهای ایجاد شده توسط آنزیمهای فوق مکمل بوده و قبلیت شناسائی و اتصال به یکدیگر را دارند. این ویژگی از اهمیت بالایی در مطالعه و دست ورزی ژنها برخوردار است.
آنزیمهای محدودکننده به دو صورت برش ایجاد میکنند. در یک حالت، آنزیم قطعاتی ایجاد میکند که در دو انتها تک رشتهای و چسبنده میباشند. برای مثال، آنزیم EcoRI قطعاتی با انتهای چسبنده ایجاد مینماید. علت چسبنده نامیده شدن دو انتهای ایجاد شده، مکملبودن آنها و قابلیت شناسائی و اتصال مجدد آنها به یکدیگر است. ناحیه چسبنده میتواند در انتهای َ۵ یا َ ۳ ایجاد شود. در حالت دوم، آنزیم دو رشته DNA را بصورت عمودی برش داده و در نتیجه قطعاتی با انتهای صاف تولید میکند. برای مثال، آنزیم HpaI با برش DNA قطعاتی با انتهای صاف ایجاد مینماید.
آنزیمهای محدودکننده نوع III
این آنزیمها از نظر عملکرد مشابه آنزیم های نوع I میباشند اما با آنکه به ATP به عنوان کوفاکتور نیاز دارند، آنرا هیدرولیز نمیکنند. EcoP۱۵(جایگاه (CAGCAG و EcoP۱ ازآنزیمهای محدودکننده نوع III میباشند.